
Effect of 40 and 80 Years of Conifer Regrowth on Soil Microbial Activities and Community Structure in Subtropical Low Mountain Forests


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Soil Sampling and Analysis
2.3. Biochemical Assays
2.4. Statistical Analysis
3. Results
3.1. Chemical and Physical Properties of Soils
3.2. Soil Microbial Biomass
3.3. Soil Enzyme Activities
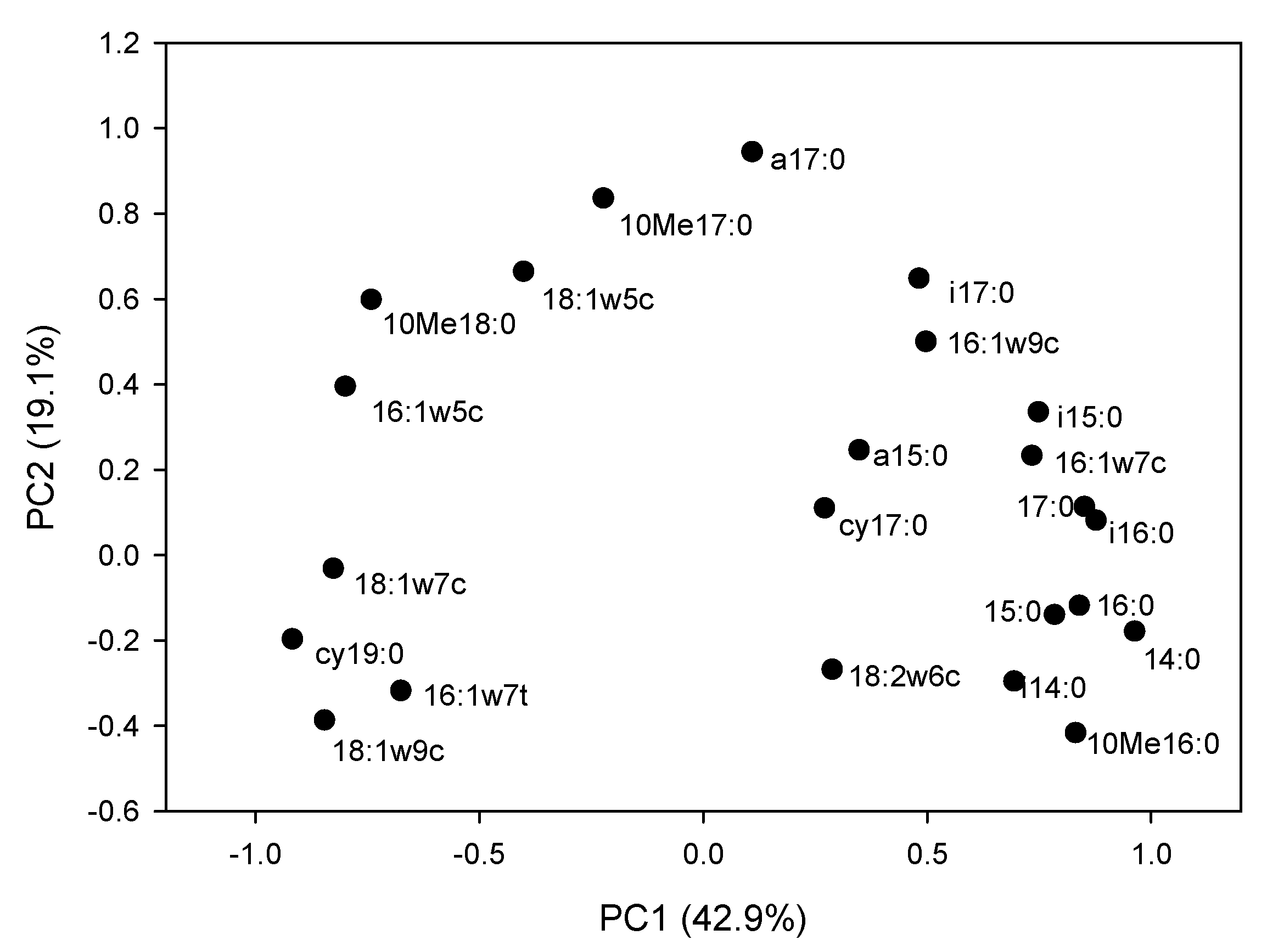
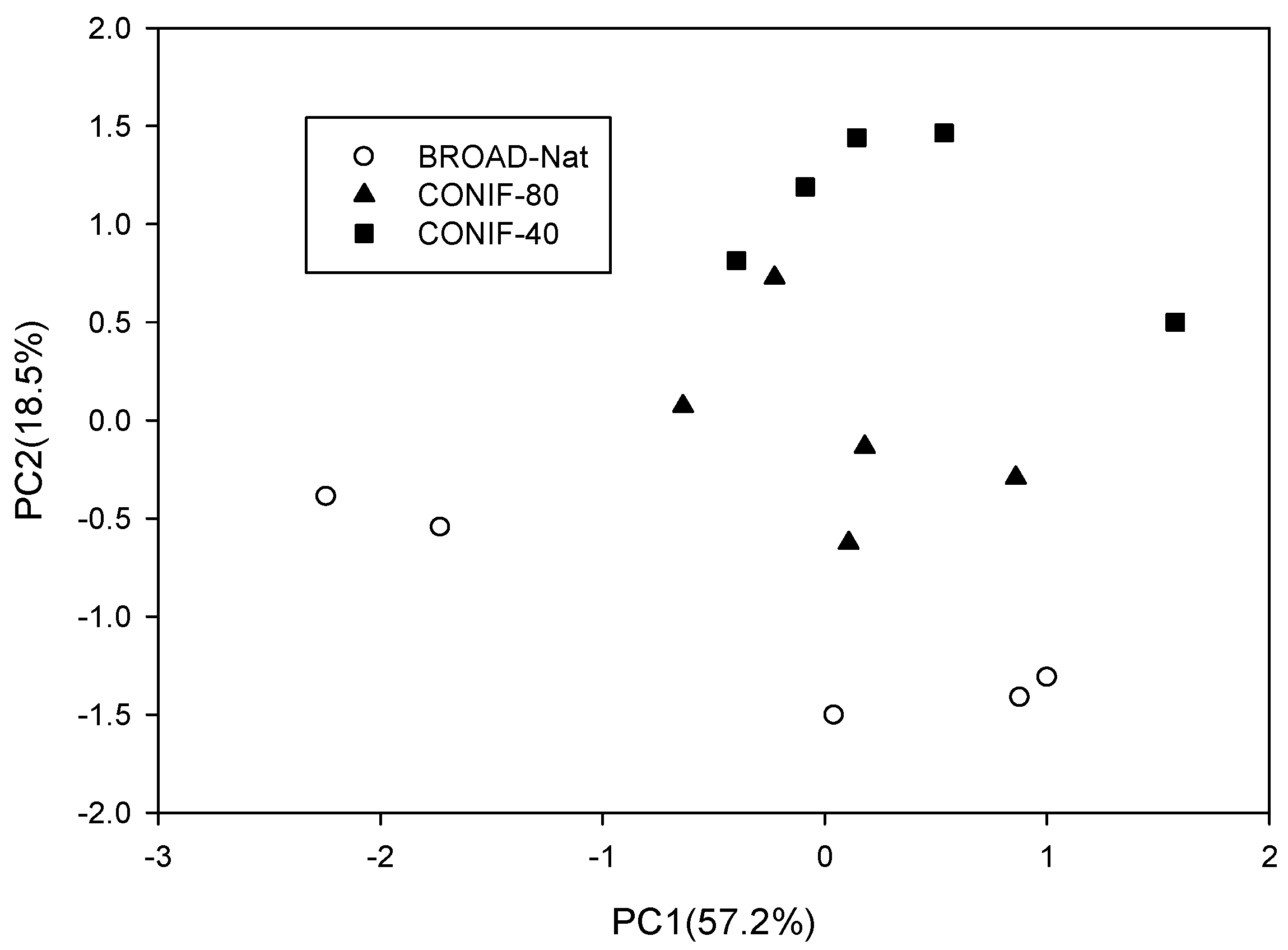
3.4. Microbial Community Structure
3.5. Correlations
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Iovieno, P.; Alfani, A.; Baath, E. Soil microbial community structure and biomass as affected by Pinus pinea plantation in two Mediterranean areas. Appl. Soil Ecol. 2010, 45, 56–63. [Google Scholar] [CrossRef]
- Chodak, M.; Niklińska, M. The effect of different tree species on the chemical and microbial properties of the reclaimed mine soils. Biol. Fertil. Soils 2010, 46, 555–566. [Google Scholar] [CrossRef]
- Samonil, P.; Král, K.; Hort, L. The role of tree uprooting in soil formation: A critical literature review. Geoderma 2010, 157, 65–79. [Google Scholar] [CrossRef]
- Banning, N.C.; Gleeson, D.B.; Grigg, A.H.; Grant, C.D.; Andersen, G.L.; Brodie, E.L.; Murphy, D.V. Soil microbial community successional patterns during forest ecosystem restoration. Appl. Environ. Microbiol. 2011, 7, 6158–6164. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Ouyang, Z.Y.; Wang, X.K.; Fang, Z.G.; Zhao, T.Q.; Miao, H. Effects of regenerating forest cover on soil microbial communities: A case study in hilly red soil region, Southern China. For. Ecol. Manag. 2005, 217, 244–254. [Google Scholar] [CrossRef]
- Cao, Y.S.; Fu, S.L.; Zou, X.M.; Cao, H.L.; Shao, Y.H.; Zhou, L.X. Soil microbial community composition under Eucalyptus plantations of different age in subtropical China. Eur. J. Soil Biol. 2010, 46, 128–135. [Google Scholar] [CrossRef]
- Ushio, M.; Wagai, R.; Balser, T.C.; Kitayama, K. Variations in the soil microbial community composition of tropical montane forest ecosystem: Does tree species matter? Soil Biol. Biochem. 2008, 40, 2699–2702. [Google Scholar] [CrossRef]
- Moore-Kucera, J.; Dick, R.P. PLFA profiling of microbial community structure and seasonal shifts in soils of a Douglas-fir chronosequence. Microb. Ecol. 2008, 55, 500–511. [Google Scholar] [CrossRef] [PubMed]
- Lou, X.; Cui, X.Y. The study of soil microbial diversity in temperate forest secondary succession process. Chin. J. Soil Sci. 2011, 3, 010. [Google Scholar]
- Overby, S.T.; Hart, S.C. Short-term belowground responses to thinning and burning treatments in southwestern Ponderosa Pine Forest of the USA. Forest 2016, 7, 45. [Google Scholar] [CrossRef]
- Chan, O.C.; Yang, X.; Fu, Y.; Feng, Z.; Sha, L.; Casper, P.; Zou, X. 16S rRNA gene analyses of bacterial community structures in the soils of evergreen broad-leaved forests in south-west China. FEMS Microbiol. Ecol. 2006, 58, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.T.; Whitman, W.B.; Coleman, D.C.; Chiu, C.Y. Molecular characterization of soil bacterial community in a perhumid, low mountain forest. Microbe. Environ. 2011, 26, 325–331. [Google Scholar] [CrossRef]
- Weand, M.P.; Arthur, M.A.; Lovett, G.M.; McCulley, R.L.; Weathers, K.C. Effects of tree species and N additions on forest floor microbial communities and extracellular enzyme activities. Soil Biol. Biochem. 2010, 42, 2161–2173. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Zhang, W.; Shao, Y.; Zou, X.; Liu, T.; Zhou, L.; Wan, S.; Rao, X.; Li, Z.; et al. Invariant community structure of soil bacteria in subtropical coniferous and broadleaved forests. Sci. Rep. 2016, 6, 19071. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.T.; Jangid, K.; Whitman, W.B.; Coleman, D.C.; Chiu, C.Y. Change in bacterial community structure in response to disturbance of natural hardwood and secondary coniferous forest soils in central Taiwan. Microb. Ecol. 2011, 61, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Zelles, L. Fatty acid patterns of phospholipids and lipopolysaccharides in the characterization of microbial communities in soil: A review. Biol. Fertil. Soils 1999, 29, 111–129. [Google Scholar] [CrossRef]
- Farrell, M.; Griffith, G.W.; Hobbs, P.J.; Perkins, W.T.; Jones, D.L. Microbial diversity and activity are increased by compost amendment of metal-contaminated soil. FEMS Microbiol. Ecol. 2010, 71, 94–105. [Google Scholar] [CrossRef] [PubMed]
- McLean, E.O. Soil pH and lime requirement. In Methods of Soil Analysis, Part 2 Chemical and Microbiological Properties; Page, A.L., Ed.; American society of Agronomy: Madison, WI, USA, 1982; pp. 199–224. [Google Scholar]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Wu, J.; Joergensen, R.G.; Pommerening, B.; Chaussod, R.; Brookes, P.C. Measurement of soil microbial biomass C by fumigation-extraction-an automated procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Amato, M.; Ladd, J.N. Assay for microbial biomass based on ninhydrin-reactive nitrogen in extracts of fumigated soils. Soil Biol. Biochem. 1988, 20, 107–114. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 8, 199–202. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Schinner, F.; Von Mersi, W. Xylanase-, CM-cellulase- and invertase activity in soil: An improved method. Soil Biol. Biochem. 1990, 22, 511–515. [Google Scholar] [CrossRef]
- Frostegard, A.; Tunlid, A.; Baath, E. Phospholipid fatty acid composition, biomass, and activity of microbial communities from two soil types exposed to different heavy metals. Appl. Environ. Microbiol. 1993, 59, 3605–3617. [Google Scholar] [PubMed]
- Chang, E.H.; Chen, C.T.; Chen, T.H.; Chiu, C.Y. Soil microbial communities and activities in sand dunes of subtropical coastal forests. Appl. Soil Ecol. 2011, 49, 256–262. [Google Scholar] [CrossRef]
- Zogg, G.P.; Zak, D.R.; Ringleberg, D.B.; MacDonald, N.W.; Pregitzer, K.S.; White, D.C. Compositional and functional shifts in microbial communities due to soil warming. Soil Sci. Soc. Am. J. 1997, 61, 475–481. [Google Scholar] [CrossRef]
- Jien, S.H.; Chen, T.H.; Chiu, C.Y. Effects of afforestation on soil organic matter characteristics under subtropical forests with low elevation. J. For. Res. 2011, 16, 275–283. [Google Scholar] [CrossRef]
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; Blackwell Scientific: Oxford, UK, 1979. [Google Scholar]
- Liu, L.; Gundersen, P.; Zhang, T.; Mo, J.M. Effects of phosphorus addition on soil microbial biomass and community composition in three forest types in tropical China. Soil Biol. Biochem. 2012, 44, 31–38. [Google Scholar] [CrossRef]
- Nannipieri, P.; Muccini, L.; Ciardi, C. Microbial biomass and enzyme activities: Production and persistence. Soil Biol. Biochem. 1983, 15, 679–685. [Google Scholar] [CrossRef]
- Badiane, N.N.Y.; Chotte, J.L.; Pate, E.; Masse, D.; Rouland, C. Use of soil enzyme activities to monitor soil quality in natural and improved fallows in semi-arid tropical regions. Appl. Soil Ecol. 2001, 18, 229–238. [Google Scholar] [CrossRef]
- Yang, Y.S.; Guo, J.F.; Chen, G.S.; Xie, J.S.; Cai, L.P.; Lin, P. Litterfall, nutrient return, and leaf-litter decomposition in four plantations compared with a natural forest in subtropical China. Ann. For. Sci. 2004, 61, 465–476. [Google Scholar] [CrossRef]
- Safari, S.A.A.; Sharifi, Z. Factors affecting cellulase activity in some soils in Hamadan Province, Iran. Iran J. Agric. Sci. 2006, 37, 645–652. [Google Scholar]
- MacKenzie, M.D.; Quideau, S.A. Microbial community structure and nutrient availability in oil sands reclaimed boreal soils. Appl. Soil Ecol. 2010, 44, 32–41. [Google Scholar] [CrossRef]
- Kytöviita, M.M.; Pietikäinen, A.; Fritze, H. Soil microbial and plant responses to the absence of plant cover and monoculturing in low arctic meadows. Appl. Soil Ecol. 2011, 48, 142–151. [Google Scholar] [CrossRef]
- Zelles, L.; Bai, Q.Y.; Beck, T.; Beese, F. Signature fatty acids in phospholipids and lipopolysaccharides as indicators of microbial biomass and community structure in agricultural soils. Soil Biol. Biochem. 1992, 24, 317–332. [Google Scholar] [CrossRef]
- Bossio, D.A.; Scow, K.M.; Gunapala, N.; Graham, K.J. Determinants of soil microbial communities: Effects of agricultural management, season, and soil type on phospholipid fatty acid profiles. Microb. Ecol. 1998, 36, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.J.; Favila, E.; Hundal, L.S.; Marlin, J.C. Assessment of soil microbial communities in surface applied mixtures of Illinois River sediments and biosolids. Appl. Soil Ecol. 2007, 36, 176–183. [Google Scholar] [CrossRef]
- Ushio, M.; Kitayama, K.; Balser, T.C. Tree species-mediated spatial patchiness of the composition of microbial community and physicochemical properties in the topsoils of a tropical montane forest. Soil Biol. Biochem. 2010, 42, 1588–1595. [Google Scholar] [CrossRef]
- Wilkinson, S.C.; Anderson, J.M.; Scardelis, S.P.; Tisiafouli, M.; Taylor, A.; Wolters, V. PLFA profiles of microbial communities in decomposing conifer litters subject to moisture stress. Soil Biol. Biochem. 2002, 34, 189–200. [Google Scholar] [CrossRef]
- Schimel, J.P.; Balser, T.C.; Wallenstein, M. Microbial stress-response physiology and its implications for ecosystem function. Ecology 2007, 86, 1386–1394. [Google Scholar] [CrossRef]
- Grayston, S.J.; Campbell, C.D.; Bardgett, R.D.; Mawdsley, J.L.; Clegg, C.D.; Ritz, K.; Griffiths, B.S.; Rodwell, J.S.; Edwards, S.J.; Davies, W.J.; et al. Assessing shifts in microbial community structure across a range of grasslands of differing management intensity using CLPP, PLFA and community DNA techniques. Appl. Soil Ecol. 2004, 25, 63–84. [Google Scholar] [CrossRef]
- Grayston, S.J.; Prescott, C.E. Microbial communities in forest floors under four tree species in coastal British Columbia. Soil Biol. Biochem. 2005, 37, 1157–1167. [Google Scholar] [CrossRef]
- Scalbert, A. Antimicrobial properties of tannins. Phytochemistry 1991, 30, 3875–3883. [Google Scholar] [CrossRef]
- Olsson, P.A.; Baath, E.; Jakobsen, I.; Soderstrom, B. The use of phospholipid and neutral lipid fatty acids to estimate biomass of arbuscular mycorrhizal fungi in soil. Mycol. Res. 1995, 99, 623–629. [Google Scholar] [CrossRef]
- Kourtev, P.S.; Ehrenfeld, J.G.; Haggblom, M. Experimental analysis of the effect of exotic and native plant species on the structure and function of soil microbial communities. Soil Biol. Biochem. 2003, 35, 895–905. [Google Scholar] [CrossRef]
- Priha, O.; Grayston, S.J.; Hiukka, R.; Pennanen, T.; Smolander, A. Microbial community structure and characteristics of the organic matter in soils under Pinus sylvestris, Picea abies, and Betula pendula at two forest sites. Biol. Fertil. Soils 2001, 33, 17–24. [Google Scholar] [CrossRef]
- Djukic, I.; Zehetner, F.; Mentler, A.; Gerzabek, M.H. Microbial community composition and activity in different Alpine vegetation zones. Soil Biol. Biochem. 2010, 42, 155–161. [Google Scholar] [CrossRef]
- Olsson, P.A. Signature fatty acids provide tools for determination of the distribution and interactions of mycorrhizal fungi in soil. FEMS Microbiol. Ecol. 1999, 29, 303–310. [Google Scholar] [CrossRef]
- Priha, O.; Grayston, S.J.; Pennanen, T.; Smolander, A. Microbial activities related to C and N cycling and microbial community structure in the rhizospheres of Pinus sylvestris, Picea abies and Betula pendula seedlings in an organic and mineral soil. FEMS Microbiol. Ecol. 1999, 30, 187–199. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest type | Species | Density (stems∙ha−1) | Plant Height (m) | Average DBH 1 (cm) | Average Basal Area (m2∙ha−1) | Wood Volume (m3∙ha−1) |
|---|---|---|---|---|---|---|
| BROAD-Nat | Cryptocarya chinensis (Hance) Hemsl. | 250 | 10.9 | 15.1 | 7.89 | 77.28 |
| Litsea acuminatae (Blume) Kurata | 175 | 12.5 | 14.7 | 4.74 | 41.85 | |
| Prunus phaeosticta (Hance) Maxim. | 175 | 13.2 | 16.7 | 4.64 | 36.20 | |
| Lithocarpus amygdalifolius (Skan) Hayata | 100 | 17.5 | 18.5 | 3.84 | 33.20 | |
| Schefflera octophylla (Lour.) Harms | 100 | 13.5 | 16.8 | 2.56 | 16.45 | |
| Cinnamomum subavenium Miq. | 175 | 13.1 | 10.5 | 2.44 | 18.44 | |
| Cyclobalanopsis pachyloma (O. Seem.) Schott. | 75 | 10.7 | 20.3 | 2.44 | 19.17 | |
| Styrax suberifolia Hook. & Arn. | 75 | 16.3 | 17.2 | 1.98 | 16.44 | |
| Pasania harlandii (Hance) Oerst. | 25 | 16.0 | 31.1 | 1.90 | 13.75 | |
| Machilus zuihoensis Hayata | 50 | 18.0 | 20.4 | 1.85 | 16.20 | |
| Others | 250 | 9.5 | 9.1 | 1.92 | 9.35 | |
| Total | 1450 | 36.20 | 298.3 | |||
| CONIF-80 | Calocedrus formosana (Florin) Florin | 275 | 26.6 | 48.0 | 51.44 | 644.9 |
| Randia cochinchinensis (Lour.) Merr | 625 | 8.6 | 6.2 | 1.95 | 7.80 | |
| Schefflera octophylla (Lour.) Harms | 575 | 8.7 | 6.2 | 1.79 | 7.24 | |
| Prunus phaeosticta (Hance) Maxim. | 275 | 10.2 | 7.5 | 1.26 | 5.96 | |
| Others | 375 | 8.0 | 5.9 | 1.15 | 4.39 | |
| Total | 2125 | 57.59 | 670.3 | |||
| CONIF-40 | Cunninghamia lanceolata (Lamb.) Hook. | 900 | 15.8 | 21.4 | 36.01 | 289.6 |
| Styrax formosana Matsum | 80 | 10.6 | 10.7 | 0.79 | 4.14 | |
| Schefflera octophylla (Lour.) Harms | 40 | 12.5 | 15.1 | 0.75 | 4.53 | |
| Others | 140 | 12.0 | 10.6 | 1.62 | 8.75 | |
| Total | 1160 | 39.17 | 307.0 |
| Season | Forest Type | Soil Moisture (%) | pH | Corg 1 (%) | Ntot (%) | Cmic (mg∙kg−1) | Nmic (mg∙kg−1) | Cmic/Corg (%) | Nmic/Ntot (%) |
|---|---|---|---|---|---|---|---|---|---|
| Summer | BROAD-Nat | 27.6 a 2 | 3.78 b | 3.45 b | 0.29 ab | 1668 a | 140 a | 4.83 a | 4.82 b |
| CONIF-80 | 23.1 b | 3.74 b | 3.23 bc | 0.24 bc | 1314 b | 94.5 c | 4.21 a | 3.93 c | |
| CONIF-40 | 29.2 a | 3.62 c | 4.45 a | 0.31 a | 1229 b | 116 b | 2.82 b | 3.74 c | |
| Winter | BROAD-Nat | 19.3 c | 3.87 a | 2.62 c | 0.22 c | 763 c | 119 b | 2.92 b | 5.40 a |
| CONIF-80 | 21.9 c | 3.87 a | 3.31 bc | 0.25 bc | 748 c | 121 ab | 2.28 c | 4.84 b | |
| CONIF-40 | 27.3 a | 3.75 b | 3.79 b | 0.29 ab | 812 c | 117 b | 2.16 c | 3.98 c | |
| ANOVA results 3 | |||||||||
| Forest type | *** | *** | ** | ** | *** | *** | *** | *** | |
| Sample season | *** | *** | ns | * | *** | ns | *** | *** | |
| Forest × season | *** | ns | ns | * | *** | *** | ns | ns | |
| Season | Forest Type | Cellulase (µg glucose g−1∙h−1) | Xylanase (µg glucose g−1∙h−1) | Urease (mmol NH4+-N g−1∙h−1) | Acid Phosphatase (µg nitrophenol g−1∙h−1) |
|---|---|---|---|---|---|
| Summer | BROAD-Nat | 31.7 c 1 | 65.8 d | 10.6 a | 569 b |
| CONIF-80 | 63.8 bc | 70.8 cd | 11.9 a | 628 b | |
| CONIF-40 | 126 a | 109 ab | 9.47 ab | 850 a | |
| Winter | BROAD-Nat | 36.7 bc | 116 a | 5.83 b | 338 c |
| CONIF-80 | 46.7 bc | 92.1 bc | 9.84 ab | 318 c | |
| CONIF-40 | 76.2 b | 118 a | 6.45 b | 361 c | |
| ANOVA results | |||||
| Forest type | ** | * | * | ns | |
| Sample Season | ns | *** | ** | *** | |
| Forest × season | ns | ns | Ns | ns | |
| Season | Forest Type | Total PLFA 1 | Bacteria | Fungi | VAM Fungi | Actino-mycetes | G+ | G− | G+/G− | Fungi/ Bacteria |
|---|---|---|---|---|---|---|---|---|---|---|
| Summer | BROAD-Nat | 115 a 2 | 50.4 a | 1.92 a | 2.59 a | 3.53 a | 23.4 a | 34.1 a | 0.69 c | 0.038 b |
| CONIF-80 | 80.7 b | 34.5 b | 1.63 a | 1.68 b | 1.99 b | 16.6 b | 23.4 b | 0.71 c | 0.047 ab | |
| CONIF-40 | 71.5 bc | 29.8 b | 1.59 a | 1.38 b | 1.80 b | 17.0 b | 16.4 c | 1.16 b | 0.052 a | |
| Winter | BROAD-Nat | 50.7 d | 12.5 c | 0.270 b | 0.611 c | 0.781 c | 6.92 d | 5.24 d | 1.32 ab | 0.021 c |
| CONIF-80 | 58.7 d | 17.4 c | 0.420 b | 0.650 c | 0.790 c | 9.80 c | 7.14 d | 1.37 ab | 0.024 c | |
| CONIF-40 | 64.1 d | 18.4 c | 0.701 b | 0.705 c | 0.777 c | 10.7 c | 7.02 d | 1.53 a | 0.038 b | |
| ANOVA results 3 | ||||||||||
| Forest type | * | * | ns | ** | *** | * | ** | *** | *** | |
| Sampling season | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| Forest × season | ** | ** | ns | *** | *** | *** | *** | ns | ns | |
| Soil Properties | Cellulase | Xylanase | Urease | Phosphatase | Total PLFA | G+ | G− | G+/G− | Fungi | VAM Fungi | Fungi/ Bacteria |
|---|---|---|---|---|---|---|---|---|---|---|---|
| C | 0.643 *** | 0.292 | 0.366 | 0.454 * | 0.320 | 0.376 | 0.177 | −0.050 | 0.132 | 0.186 | 0.159 |
| N | 0.476 ** | 0.118 | 0.406 * | 0.394 * | 0.464 * | 0.498 ** | 0.325 | −0.050 | 0.132 | 0.339 | 0.206 |
| Cmic | −0.039 | −0.606 *** | 0.503 ** | 0.619 *** | 0.895 *** | 0.917 *** | 0.964 *** | −0.902 *** | −0.651 *** | 0.952 *** | 0.888 *** |
| Cmic/Corg | −0.365 | −0.753 ** | 0.268 | 0.363 | 0.711 *** | 0.711 *** | 0.843 *** | −0.884 *** | −0.672 *** | 0.821 *** | 0.776 *** |
| Cmic/Nmic | 0.112 | −0.584 ** | 0.459 * | 0.676 *** | 0.665 *** | 0.743 *** | 0.819 *** | −0.889 *** | 0.670 *** | 0.779 *** | 0.919 *** |
| pH | −0.678 *** | −0.148 | −0.273 | −0.682 *** | −0.419 * | −0.513 ** | −0.381 * | 0.247 | 0.202 | −0.389 * | −0.492 ** |
| Moisture | 0.428 * | −0.039 | 0.291 | 0.529 ** | 0.637 *** | 0.680 ** | 0.529 ** | −0.252 | −0.023 | 0.556 ** | 0.440 * |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, E.-H.; Chen, T.-H.; Tian, G.-l.; Hsu, C.-K.; Chiu, C.-Y. Effect of 40 and 80 Years of Conifer Regrowth on Soil Microbial Activities and Community Structure in Subtropical Low Mountain Forests. Forests 2016, 7, 244. https://doi.org/10.3390/f7100244
Chang E-H, Chen T-H, Tian G-l, Hsu C-K, Chiu C-Y. Effect of 40 and 80 Years of Conifer Regrowth on Soil Microbial Activities and Community Structure in Subtropical Low Mountain Forests. Forests. 2016; 7(10):244. https://doi.org/10.3390/f7100244
Chicago/Turabian StyleChang, Ed-Haun, Tsai-Huei Chen, Guang-long Tian, Chun-Kai Hsu, and Chih-Yu Chiu. 2016. "Effect of 40 and 80 Years of Conifer Regrowth on Soil Microbial Activities and Community Structure in Subtropical Low Mountain Forests" Forests 7, no. 10: 244. https://doi.org/10.3390/f7100244