3.1. Reduction to the Uncontrolled System and Basic Dynamical System Properties
We carry out the dynamical systems analysis for constant controls, i.e., concentrations. We do not explicitly include pharmacokinetics (fluctuations in concentrations that depend on doses). The treatments considered are either administered daily or have long half-lives, and such pharmacokinetics are not expected to be significant for the treatment periods we consider here (five years or longer). We also mention the 2009 paper by Shudo et al. [
16] that supports this assumption in the setting of hepatitis C.
Keeping the “
” parameters in their original formulation in the controls, we define new drug-dependent parameters as
With these identifications, the dynamical system with constant controls is identical with the uncontrolled system and therefore, without loss of generality, the analysis can be done on the uncontrolled system. Returning to the original notation without the carets, we thus consider the following equations:
The model with an exponential growth term on
Q has various long-term behaviors. These include the extremes in which
Q decays exponentially to zero or grows exponentially beyond limits, but there also is the possibility that nontrivial equilibrium points
exist for which all three populations are positive. The first case corresponds to a scenario in which the patient goes into a stable deep molecular response. For the uncontrolled system, this may not seem to be of interest, but since the model includes the case with controls, this gives us information about which combinations of constant concentrations of the drugs would lead to an eradication of
Q. The case of exponential growth may characterize the
accelerated or
blast phase as these phases have short doubling times [
17]. The conditions under which this is the long-term behavior of the system give information about what controls are needed for successful treatment. An asymptotically stable equilibrium point
with positive values could be interpreted as describing a subset of the
chronic phase where net growth rate is zero, controlled by therapy or immune effects. Depending on the values of the parameters, this equilibrium point may be stable or unstable. Since in real life parameters may not be constant, bifurcation phenomena would be a mathematical description of the transition from chronic to the accelerated or blast phases. Knowing the parameter values when this may occur would be of interest. Our aim in the following is thus to determine the asymptotic behavior of the trajectories of the system.
We start with some basic properties. The positive orthant
is positively invariant for the dynamics. This is because the planes
and
are invariant under Equations (
6) and (
8) and
whenever
. Thus, starting at a positive initial condition
, it follows that the solutions remain positive for all times. For the long-term behavior of the system, the equilibrium solutions in the closure of
,
, also matter. Recall that the system is defined and continuous on
due to the use of the limits as
and
in place of
and
, respectively. The vector field defining the
P and
E dynamics is not continuously differentiable at
or
, but these values are repelling and thus this does not become an issue.
Lemma 1. The equilibrium solution is repelling: there exists a positive threshold such that is positive on . In particular, once , then for all . Furthermore, for , E will remain below .
Proof. The terms in the last parentheses in Equation (
8) are bounded between 1 and
and thus, as
, the Gompertzian growth dominates the dynamics. Specifically, let
then
and for
we have that
. Furthermore, for
, Equation (
8) reduces to
and thus the values of
E cannot reach the value
if they start below
.
Lemma 2. The equilibrium solution is repelling: there exists a positive threshold such that is positive on . In particular, once , we have for all .
Proof. For values of
E less than
, we have that
for all times. Choosing
as
the result follows: for
we have that
This proves the result.
Corollary 1. The equilibrium solutions and are unstable.
Note, however, that
P is not necessarily bounded. For, with
, Equation (
7) becomes
and thus, if
Q is large enough, this term will be positive. Hence, if
Q grows exponentially,
P will diverge to
.
Lemma 3. If Q increases exponentially with time, then .
Proof. We need to show that for every positive value there exists a time so that for all .
We first remark that
P is unbounded. For, if there exists a value
with
so that
for all times
t, then the term
is bounded below. By assumption, there exist positive constants
α and
β so that
for all
t. Hence, for
t sufficiently large we have that
Contradiction.
Given
, choose
so that
Since
P is not bounded, there exists a first time
so that
. We claim that
for all
. For, if there exists a time
such that
, then
Contradiction. Thus P diverges to .
3.2. Dynamics on the Plane
The plane
is invariant under the dynamics and can have regions that are repelling or attractive. We first analyze the reduced dynamical system in this boundary stratum of
, i.e., consider the equations
Let
denote the open rectangle
and denote by
its closure,
. For
the variable
P is bounded above by
and therefore the compact set
is positively invariant under Equations (
9) and (
11). The dynamical system has the following
trivial equilibrium solutions in the boundary of
:
,
with
and
with
given by
In view of Lemmas 1 and 2 these solutions are unstable. While the origin has two unstable modes, the equilibrium points and are saddles with the respective axes forming the stable manifolds and the unstable modes entering the interior of . It is clear from this that there needs to exist at least one more equilibrium point in .
Lemma 4. There are no periodic orbits in .
Proof. Changing variables to
and
, the dynamics transforms into
The divergence of this vector field is given by
and thus the result follows from Bendixson’s negative criterion because of the monotonicity of the logarithm function.
The relations defining equilibrium points inside
are
and
or, equivalently,
Then equilibrium values
are fixed points of the function Φ,
, in the interval
. Since
,
, and Φ is continuous in
P, it follows that there exists at least one solution. The derivative
of Φ is given by
and thus has the same sign as
. Now
Thus Φ is strictly increasing for
and strictly decreasing for
. If
, then
Equilibria are intersections of the graph of Φ with the diagonal and thus there exists a unique equilibrium point if , but multiple solutions are possible if .
We determine the stability of
for the reduced system, i.e., within the invariant plane
. The Jacobian matrix at the equilibrium point is given by
Using the equilibrium relations we can write the
-term as
The characteristic polynomial of this
matrix is given by
If we write , then is positive and thus the equilibrium point is locally asymptotically stable if is positive while it is unstable if is negative. A saddle node bifurcation occurs as . It immediately follows that is locally asymptotically stable if , i.e., if the stimulating effect of the tumor on the effector cells is larger than the inhibiting effect of the tumor on the effector cells. We have the following result:
Proposition 1. If , then there exists a unique equilibrium point in and it is globally asymptotically stable in the sense that its region of attraction is the full rectangle .
Proof. The set is positively invariant and every trajectory γ starting in has a non-empty ω-limit set . Because of the stability properties of the equilibria in the boundary of , this ω-limit set lies in . Since there exist no periodic orbits and since is the only equilibrium point, it follows from Poincaré-Bendixson theory that , i.e., all trajectories starting in converge to as . ☐
It is clear from Poincaré-Bendixson theory that even if
, the equilibrium point
is globally asymptotically stable (in the sense that its region of attraction contains the set
, and only this region is relevant for the problem) as long as it is the only equilibrium point in
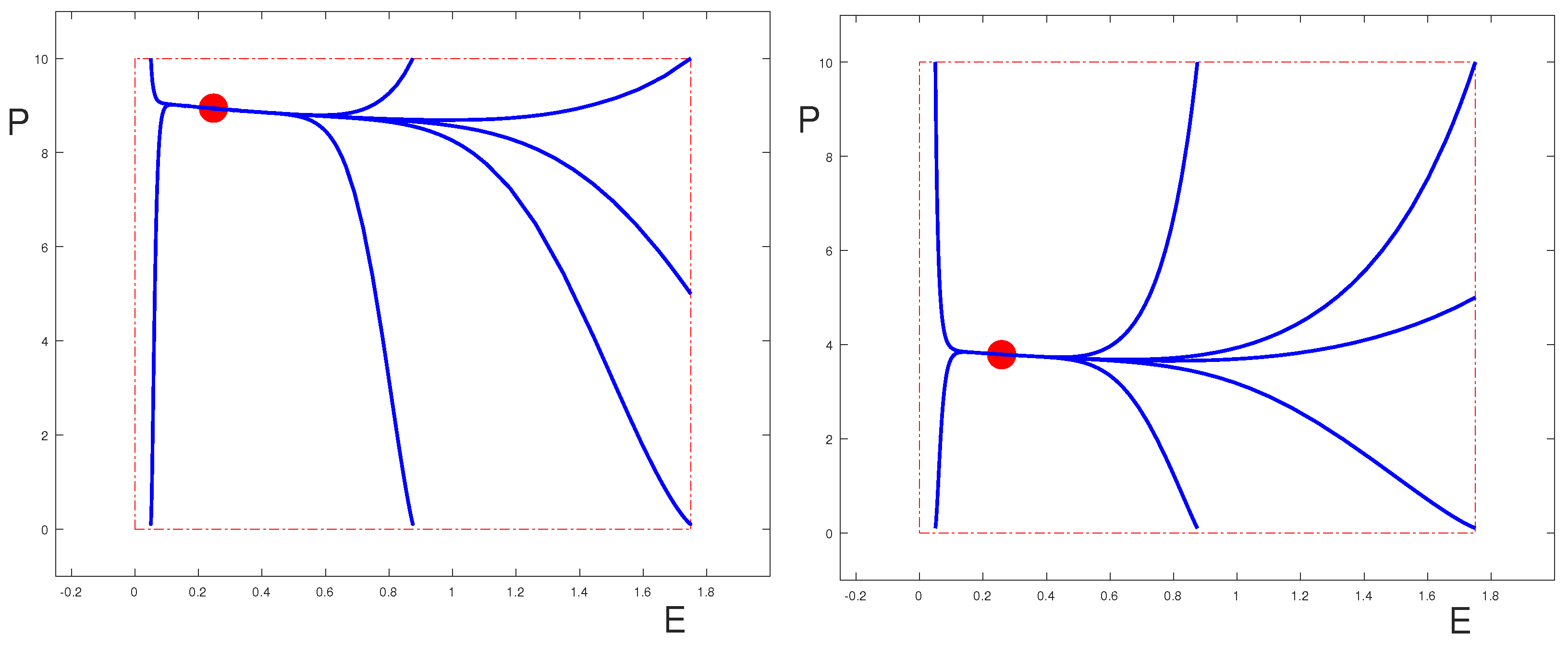
. This is shown in the phase portraits for the uncontrolled system in
Figure 2;
Figure 3 shows a case where
. (The values of the parameters are given in
Table 1 and
Table 2.) We also show the phase-portraits for the systems when one of the controls is set to be equal to 1 and all others are zero. The two sets of figures illustrate two different scenarios, one where the control parameters are such that the equilibrium can be effectively controlled by all the drugs (
Figure 2), the other where it is essentially only the control
that is able to move the equilibrium point. However, this behavior depends on the fact that
.
Since the coefficient is always positive, as vanishes the Jacobian matrix has the eigenvalue 0 and the other eigenvalue is negative. At such a point saddle-node bifurcations arise and two new equilibria, one stable, the other unstable, are born.
Proposition 2. If , then multiple equilibria inside can exist. At points wheresaddle-node bifurcations occur in which a stable and an unstable equilibrium point merge. Proof. The coefficient
vanishes if and only if
This condition is equivalent to (
14).
For the underlying biological problem it is natural that an inhibition effect would be smaller than a stimulation effect. Also, the denominators are quadratic in the respective variables
E and
P, but these variables are scaled. In principle it is possible to satisfy (
14), but we did not come across this in our simulations.
3.3. Dynamic Behavior for Positive Q-Values
For the behavior of the overall system, the
Q dynamics are essential. If one considers the above equilibria in the plane
now in the full three-dimensional space, then the first row of the Jacobian matrix at
takes the form
and thus
is unstable if
while the local stability properties for the overall system are the same as in the
-plane if
. If
, then there exists a 1-dimensional center manifold (corresponding to the 0 eigenvalue). In this case we have
and there exists a curve of equilibria emerging from
parameterized by
Q or
P (also see below).
Generally, (
1)–(
3) is a time-varying linear system dominated by exponential growth and decay, depending on the parameter values. If
then
Q grows exponentially and no steady state exists. In this case, the influx
eventually becomes the dominant term in Equation (
2) and
P also grows beyond limits (Lemma 3). This represents the
malignant scenario in the model which corresponds to a highly-aggressive form of the disease or the
accelerated or
blast phase. The other extreme arises if
. In this case
Q exponentially decays to 0 for the uncontrolled system and overall trajectories converge to one of the equilibria
in the plane
. If there exist multiple such equilibria, there exists a stable manifold for the unstable one that separates the regions of attraction for the stable equilibria. This would reflect a scenario when
Q initiates the disease, but eventually dies off and the remaining
P population determines the outcome of the disease. This could be benign if
is small (a form of successful
immune surveillance) or malignant if this value is larger. In such a case, however, one only needs to deal with the proliferating cells as far as treatment is concerned. This appears less likely (unless it could be induced by the drugs) and in the uncontrolled case of the disease we would have
.
The interesting and most difficult case arises when the uncontrolled system has a chronic steady state or undergoes exponential growth without treatment, but has a negative net growth rate for
Q with treatment. This is the case if the parameters satisfy the following condition (A):
or, with the controls in the original form,
Thus the replication rate constant
needs to be greater than the death rate constant
(this naturally will be satisfied for parameters in a disease state), but at the same time, the drugs need to be able to raise the maximum effectiveness
high enough that the magnitude of the immune system effect can overcome the difference. These appear to be natural conditions. Assuming that (
15) holds, there exists a unique value
for which
, namely
with
Q increasing for
and decreasing for
. In this case, the interplay between the variables allows for a steady state
to exist with all values positive. We call such an equilibrium point
positive.
3.5. Existence and Stability of a Positive Equilibrium Point for
We analyze whether positive equilibrium points
exist. Throughout this section we assume that condition (
15) is satisfied, i.e., that
since otherwise
Q grows exponentially.
Lemma 5. For , there exists at most one positive equilibrium point .
Proof. The equilibrium relation for Equation (
6) uniquely determines
:
Given
, the equilibrium condition on the effector cells,
, is equivalent to
The quantity
is already determined. If
, then (
17) only has the solution
; otherwise there exists a unique solution
given by
If
, this solution is positive if and only if
and if
, the solution is positive if and only if
If one of these inequalities is violated, no positive equilibrium solution
exists and the overall dynamics are determined either by exponential growth or decay of
Q. If
exists and is positive, then Equation (
7) defines
as
Using the equilibrium relation for
, this can equivalently be expressed in the form
Note that
is positive if
while otherwise this becomes a requirement on the equilibrium value
, namely
If
, then this simply becomes
. In either case, there exists at most one positive equilibrium point given by Equations (
16), (
18) and (
20).
Remark 1. As , condition (15) implies that along a positive solution we must haveand thus the limit taken along these positive solutions only exists if and if In this degenerate case, the equilibrium conditions and are automatically satisfied and there exists a one-dimensional equilibrium manifold, namely with the P value arbitrary and given by We now investigate the
stability of the positive equilibrium point. The partial derivatives of the equations defining the dynamics at the equilibrium point are given by
Note that the equilibrium condition for
P brings in
in
. The characteristic polynomial for the Jacobian matrix is given by
By the Routh-Hurwitz criterion, all eigenvalues have negative real parts if and only if
,
and
. These coefficients are given by
If , then is negative and the positive equilibrium point is unstable, i.e., once the maximal inhibiting effect of the tumor on the effector cells is larger than the maximal stimulating effect, no steady-state positive solution exists. Note further that for the characteristic polynomial has exactly one change of sign in its coefficients and thus there exists a unique positive root. So the equilibrium point has a two-dimensional stable manifold that separates the regions where Q and P diverge to infinity from the region where Q converges to 0. Thus we have the following result:
Theorem 1. If , then the positive equilibrium point is unstable with a two-dimensional stable manifold in parameter space.
If , then the equilibrium point has the eigenvalue 0 and two negative eigenvalues. Thus there exists a one-dimensional center manifold which in this case consists of all equilibria, namely the equilibrium manifold M defined earlier.
For
, the coefficients
,
and
are all positive. Furthermore
This expression is positive if we make the following assumption (B):
Note from Equations (
2) and (
3) that
represents the maximal size of the immune effect
E on
P while
represents the maximal size of the immune effect
E on
Q. This effect is assumed to be stronger on the proliferating class of cells than on the quiescent class of cells. Thus assumption (
21) is a natural one to make. This assumption is invariant under the actions of the drugs:
while, letting
,
so that these terms are multiplied by the same coefficients. Hence we also have the following result:
Theorem 2. If and , then the positive equilibrium point is locally asymptotically stable.
The limiting case
represents a degenerate scenario. In many cases no positive equilibrium exists. For example, if
, then it follows from (
18) that
In such a case equilibria will cease to exist, as
, once the parameter values satisfy
Also, although the positive equilibrium point in Theorem 2 is stable, the value can be very high. In fact,
diverges to
as these parameter relations are reached (c.f. (
18)):
For the equilibrium values to be relatively small (‘chronic’), we see that
must be significantly larger than
. In terms of the parameter values with drug actions, this can be achieved using the drugs
and
which increase
and decrease
, c.f.,
So drug administration shifts the balance towards and this creates an asymptotically stable positive equilibrium point , hopefully with low values for and .
Figure 4 shows how the positive equilibrium values change as (only) one of the controls is varied. Note that the equilibrium values for
Q and
E do not change if only the control
is varied. Also for changes in the controls
and
these equilibrium values change little and in the graphs the corresponding curves are almost constant. However, in these cases the equilibrium values for
Q and
P are well-controlled by the therapies. Contrary to the case when
, the
and
controls have strong effects by cutting down the influx of cells from the
Q into the
P compartment. All equilibria shown in these graphs satisfy the conditions of Theorem 2 and are locally asymptotically stable.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}